The von Bertalanffy growth function (VBGF) introduced by von
Bertalanffy in 1938 predicts the length of a shark as a function of its
age, L = L(t):
L(t) = Loo - (Loo - Lo) exp(-kt)
1. Lo (Lzero, y-axis intercept) is the mean length at birth (t =0),
2. Loo (L infinity) is the mean maximum length (t = infinity),
3. k is a rate constant with units of reciprocal time (e.g. year-1).
The graph on the right used Lo = 0.5 m, Loo = 3 m, k = 0.13863 year-1
(ln2/k = 5 years;
4 ln2/k = 20 years;
5 ln2/k = 25 years;
7 ln2/k = 35 years)
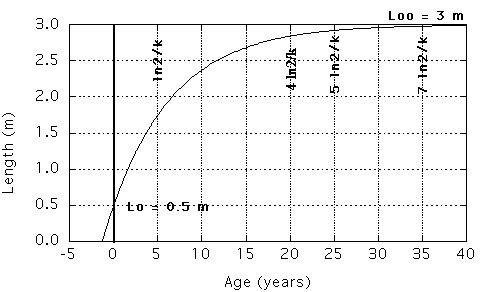
The difference between Loo and Lo diminishes ("decays") exponentially. Ln2/k is a half-life i.e. in this time the shark will be halfway between Lo and Loo. 4ln2/k, 5ln2/k and 7ln2/k are longevity estimates. In a first approximation (Lo<<Loo): In this time the shark will have reached x = 93.75, 96.875 and 99.21875 %, respectively, of the mean maximum length Loo [Correct would be to say that these %ages apply when the shark has reached Lo + (Loo-Lo)*x/100.]
The formula to calculate the time needed for a shark to have reached
the fraction x of Loo is given by
t (in units of yr) = 1/k ln[(Loo-Lo)/((1-x)Loo)]
t (in units of ln2/k) = 1/ln2 ln[(Loo-Lo)/((1-x)Loo)] = 1.443 ln[(Loo-Lo)/((1-x)Loo)].
It is easier to understand/discuss the VBGF using longevity (with units of time) instead of k (with units of time^-1), despite the fact that longevity is rather ill-defined. It also better reflects the meaning of the VBGF. Accordinng to Ricker (1979, p. 720) it was Taylor (1958) who proposed that longevity should be defined as the time when L(t) reaches 95% of Loo [L(t)/Loo = 0.95]. This could be considered to be the lower limit although Taylor proposed it to be an upper age/longevity limit. In the example above, for x = 0.95, we obtain t = (1/0.13863) ln[(3.0-0.5)/((1-0.95)3.0)]
= 20.3 years. The shark has reached 95% of Loo in 4.06 half-lives = 20.3
years.
Fabens (1965) suggested that longevity should be defined as the time when L(t) becomes larger than 99% [L(t)/Loo > 0.99, the exact value is 0.994383] with associated time of 5/k (note that 5/k = 7.213 ln(2)/k). In the example shown above this would be 36.065 yr and could be considered to be the upper limit. The shark has reached 99.4% of Loo in 7.213 half-lives = 36.1 years. Note that the upper limit (Fabens) is almost twice as large as the lower limit (Taylor)
k has often been called a growth constant. In the demonstration graph
below I used 2 pups with the same length at birth (0.5 m) and weight (0.625
kg). I used the same anabolic constant a (usually denoted as eta) and
2 different values for k (3k = catabolic constant, usually denoted as
chi) and calculated growth rates vs. age. A large k (0.25 yr-1) produces a mature
adult shark of low mass (8.64 kg) which is reached in a short time. This
may look like fast growth, however, the corresponding growth rates (with
units of kg/yr or m/yr) are small. With a smaller k of 0.1 yr-1, the maximum
growth rates are 6 x as large and the shark reaches a considerably larger
mass (135 kg), but it takes a lot longer to reach the steady state.
dM/dt = a M^(2/3) - 3k M
1. Mo = 0.625 kg;
2. a = 1.539 kg^0.333 yr-1;
3. k = 0.1 and 0.25 yr-1.
The anabolic constant a was chosen to produce Moo = 135 kg with k = 0.1, where 135 kg was the estimated mass of a shark of 3 m TL assuming M = 5 TL^3.0.
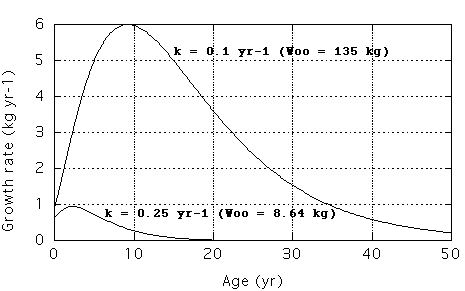
Miscellaneous items
-
As outlined above k is not a growth constant but apparently sharks
with large growth rates have large k. k (rate constant) and dL/dt (growth
rate) are related as follows:
If "growth" is understood to be the growth rates of pups or juveniles (large adult sharks no longer grow much) then we can replace L with Lo and now compare two shark of SIMILAR Loo. Furthermore, let's assume that Loo >> Lo, and we have k ~ (dL/dt)/Loo i.e. k is proportional to dL/dt.
[Based on a recent conversation with Jeff Carrier (Nov 2015) a different way to look at this is as follows: The VBGF is L(t) = Loo - (Loo - Lo) * exp (-k*t), the growth rate is then dL(t)/dt = k * (Loo - Lo) * exp (-k*t). At age 0 (t = 0, birth) we have dL/dt = k * (Loo - Lo). If we assume that (Loo - Lo) is constant or does not change considerably for different sharks in a first rough approximation, then the growth rate at birth (with units of length/time) is approximately proportinal to k (with units or reciprocal time.]k in the VBGF is assumed to be a constant. However, Von Bertalanffy (1960) indicated that k could change as a shark get older. Sevengill shark growth data indicated that indeed k is not constant and becomes smaller as they get older.
The VBGF is a 3 parameter equation. In principle, three data points determine the parameters. If adult males and females reach different maximum sizes, then the VBGF of males and females must be different because size at birth and say first year growth of males and females are the same.
Most statistical packages include a non-linear module which can be used to calculate the best fitting parameters for the available length age data pairs. Most frequently least-squares are used but a maximum likelihood loss function may produce more robust parameters.
The VBGF given here first, is the most suitable form for sharks which have a well defined size at birth. The following theoretical publications and papers dealing with elasmobranch research used this form:
Aasen, O. 1963. Length and growth of the porbeagle (Lamna
nasus, Bonnaterre) in the North West Atlantic. Rep. Norw. Fishery
Mar. Invest. 13: 20-37.
Bertalanffy, L. von 1938. A quantitative theory of organic
growth (Inquiries on growth laws. II). Human Biol. 10: 181-213.
Bertalanffy, L. von 1960. Principles and theory of growth,
pp 137-259. In Fundamental aspects of normal and malignant growth.
W. W. Wowinski ed. Elseviers, Amsterdam.
Cailliet, G. M., H. F. Mollet, G. G. Pittenger, D. Bedford, and
L. J. Natanson 1992. Growth and demography of the pacific angel
shark (Squatina californica), based upon tag returns off California.
Australian Journal of Marine and Freshwater Research 43: 1313-30.
Fabens, A. J. 1965. Properties and fitting of the von Bertalanffy
growth curve. Growth 29: 265-289.
Van Dykhuizen, G. and H. F. Mollet 1992. Growth, age estimation,
and feeding of captive sevengill sharks, Notorynchus cepedianus,
at the Monterey Bay Aquarium. In Sharks: Biology and Fisheries.
J. G. Pepperell ed. Australian Journal of Marine and Freshwater Research
43: 297-318.
The VBGF is more often presented in a different form which uses to (t zero, x-axis intercept) as the 3rd parameter rather than Lo (L zero, y-axis intercept).
t-zero was assumed to be the gestation time (time from fertilization to birth) by many (e.g. Holden 1974) but this implies that embryonic growth follows the same growth law governing post-natal growth. No data were ever produced to substantiate this, wheras available data of embryonic growth suggests that it is different from post-natal growth and thus requires its own growth curve. Indeed, it would be surprising if embryonic growth of elasmobranchs featuring a large number of reproductive modes (ovipaity to placental vivipartiy) would follow/determine) post-natal growth. Accordingly, t-zero has little meaning and I suggest that it is preferable to use Lo when reporting VBGF's of elasmobranchs. Lo can be calculated from the parameters Loo, k, and t zero from the following equation Lo = Loo[1 - exp(kto)]. The example in the graph has to = (1/k)ln[(Loo-Lo)/Loo] = 1.6 years.
The VBGF is sometimes used in yet another form involving as the third parameter b = (Loo-Lo)/Loo = exp(kto)
where T = time-at-large. The age of the shark is not required. If age-at-birth (Lo) is known a 3-paramter curve can be constructed. This does not mean that length-age data can/should be fitted to a 2-parameter VBGF with fixed (predetermined) Lo.
Growth rate vs. age (explicit age dependence)
Explicit expression of growth rate in units of length/time (e.g. m/year).
Growth rate vs. length (implicit age dependence, Gulland-Holt method)
Age not required. Annualized growth rates are plotted vs. mean length at tagging and recapture. Times-at-large don't have to be equal. If times-at large vary too much, a correction factor can be used.
Fractional growth rate L'/L = k [(Loo/L) - 1]
Often k is misnamed as a growth constant although k has units of reciprocal time. The Gulland-Holt equation shows how k and the growth rate are related: k = L'/(Loo - Lo) where L' = L'(t)
Gulland method, L increment vs. L at tagging
where T = time at large. Should be the same for all the data. Age is not required.
VBGF equations for mass, CRM = cube root mass
growth rate vs. age
fractional growth rate vs. age, related to food intake vs. age.